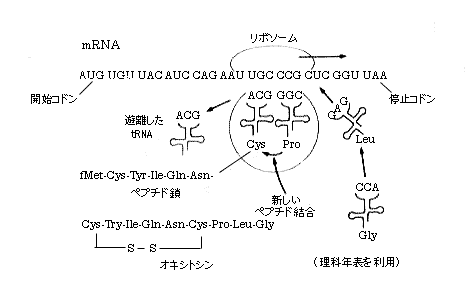
侽俀丏堚揱巕偲宍幙敪尰乮懱幙乯偲偺娭學偼丗 乽堚揱巕偼DNA暘巕偺堦椞堟傪愯傔丄摿掕偺墫婎攝楍傪桳偟丄摿掕偺宍幙傪敪尰偡傞乿 偟偨偑偭偰丄堚揱巕偺惁偝傪棟夝偡傞偨傔偵偼丄DNA暘巕偺墫婎攝楍偲宍幙敪尰偲偺娭學傪挷傋傞昁梫偑偁傞丅 偙偙偱丄DNA偲偼deoxyribonucleic acid丄僨僆僉僔儕儃妀巁偺棯徧偱偁傝丄俀庬椶偁傞妀巁乮nucleic acid乯偺侾庬偱偁傝丄巁惈崅暘巕暔幙偱偁傞丅傕偆堦偮偼RNA偱偁傝丄ribonucleic acid丄儕儃妀巁偱偁傞丅 DNA乮deoxyribonucleic acid乯偵偼丄俀偮偺廳梫側栶妱偑偁傝傑偡丅帺屓暋惢乮嵶朎暘楐偲憹怋乯偲僞儞僷僋幙崌惉偱偡丅屻幰偵傛偭偰惗惉偝傟傞僞儞僷僋幙偑丄屄恖屄恖偺堚揱宍幙傪宍惉丒巟攝偟傑偡丅 偱偼丄DNA丄墫婎攝楍丄宍幙敪尰丄偦偟偰僞儞僷僋幙崌惉偺憡屳娭學偵偮偄偰丄乮侾乯偲乮俀乯偺俀偮偵暘偗偰奣愢偟傑偡丅 乮侾乯僞儞僷僋幙偲宍幙偲偺娭學丗 変乆偺懱撪偱婲偒偰偄傞斀墳偺杦偳慡偰偵峺慺偑娭學偟偰偄傞丅斀墳偺庬椶偵墳偠偰峺慺偺庬椶傕堎側傞偑丄偙偺峺慺偼僞儞僷僋幙暘巕偱偁傞丅傑偨丄嵶嬠偑懱撪偵怤擖偟偨帪偵杊屼偡傞峈懱傕僞儞僷僋幙暘巕偱偁傞丅嵶朎偺庡梫側峔惉梫慺傕僞儞僷僋幙偱偁傝丄僀儞僔儏儕儞偺傛偆側儂儖儌儞傗愒寣媴拞偺巁慺傪塣傇僿儌僌儘價儞傕僞儞僷僋幙偱偡丅偙偺傛偆偵丄僞儞僷僋幙偑惗柦妶摦偺廳梫側埵抲傪愯傔偰偄傑偡丅堚揱巕偵寚娮偑偁傞偨傔偵婲偒傞堚揱巕昦偲偟偰丄俁丆侽侽侽庬傕偺昦婥偑偁傞偦偆偱偡丅偨偲偊偽丄姍忬愒寣媴昻寣徢偲偄偆愭揤惈幘姵乮堚揱宍幙乯偑偁傝傑偡偑偙傟偼丄DNA偺墫婎攝楍偺偨偭偨侾屄強偵堎忢偑偁傞偨傔偵丄惓忢偺応崌偲偼堎側傞僞儞僷僋幙偑崌惉偝傟傞偙偲偵傛偭偰惗偢傞堚揱揑幘姵偱偁傝傑偡丅 埲忋偐傜丄乽僞儞僷僋幙偑宍幙敪尰偵戝偒偔娭傢偭偰偄傞乿偲偄偆偙偲偑傛偔暘偐傝傑偡丅偦傟偲丄乽崌惉偝傟傞僞儞僷僋幙偺庬椶偼丄DNA偺墫婎攝楍偑寛傔傞乿偲偄偆偙偲傕憐憸偱偒傑偡丅 乮俀乯DNA偺墫婎攝楍偑僞儞僷僋幙偺庬椶傪寛傔傞丗 僸僩偺嵶朎撪偵偍偗傞僞儞僷僋幙崌惉偱偼丄愭偢DNA偺堚揱忣曬偼儊僢僙儞僕儍乕RNA偲偄偆暘巕偑崌惉乮揮幨乯偝傟丄師偄偱儊僢僙儞僕儍乕RNA偼儕儃僜乕儉偵寢崌偟丄僞儞僷僋幙惗崌惉偑峴傢傟傞丅 嘆揮幨丗 嵶朎撪晹偺妀偵偍偄偰丄DNA偺堚揱忣曬偱偁傞墫婎攝楍偼丄偄偭偨傫RNA偺墫婎攝楍偵抲偒姺偊傜傟傞丅偙偺暔幙傪儊僢僙儞僕儍乕RNA乮mRNA乯偲屇傃丄偙偺僾儘僙僗傪揮幨偲偄偆丅揮幨僾儘僙僗偵偼RNA崌惉峺慺乮RNA億儕儊儔乕僛乯偑廳梫側栶妱傪壥偨偟偰偄傞丅 RNA億儕儊儔乕僛偼丄DNA偺墫婎攝楍傪拻宆偲偟偰丄RNA傪崌惉偡傞峺慺乮偙傟傕僞儞僷僋幙偱偡乯偱偁傞丅偡側傢偪丄RNA億儕儊儔乕僛偑DNA偺偁傞晹埵偵寢崌偡傞偲丄偦偺晹埵廃曈偺DNA偺俀廳傜偣傫偼彮偟姫偒栠偝傟丄墫婎懳偑偼偢傟偰侾杮嵔DNA偲側傞丅偙偙偱丄偙偺峺慺偑嶌梡偟偰侾杮嵔DNA偲憡曗娭學偵偁傞RNA偑崌惉偝傟傞丅偙傟偑揮幨偲屇偽傟傞桼墢偱偁傞丅偦偺晹埵偱偺崌惉偑廔傞偲丄RNA億儕儊儔乕僛偼DNA忋傪儗乕儖偺傛偆偵堏摦偟丄師乆偲揮幨嶌嬈傪孞傝曉偟偰RNA傪崌惉偡傞丅偙偺揮幨埵抲偺巒傔偲廔傝傪擣幆偡傞偺傕RNA億儕儊儔乕僛偺栶妱偱偁傞丅 偙偺傛偆偵偟偰崌惉偝傟偨mRNA偼丄妀奜傊弌偰嵶朎幙偺儕儃僜乕儉偲寢崌偟丄僞儞僷僋幙崌惉斀墳偵娭梌偡傞丅偙偺僾儘僙僗傪東栿偲偄偆丅 側偍丄RNA億儕儊儔乕僛偼俁庬椶偁傝丄mRNA崌惉傪峴偆偺偼RNA億儕儊儔乕僛嘦偱偁傞丅 嘇東栿丗 妀撪偱崌惉偝傟偨mRNA偼妀奜乮嵶朎偺拞乯偵弌偰丄儕儃僜乕儉乮rRNA偲僞儞僷僋幙偺暋崌懱偱丄僞儞僷僋幙崌惉偑峴傢傟傞応強偲側傞棻巕乯傗揮堏RNA乮tRNA乯丄偦偟偰傾儈僲巁傗峺慺乮儁僾僠僕儖僩儔儞僗僼僃儔乕僛乯偺嫤椡偺壓偵丄DNA偺墫婎攝楍偵懳墳偟偨僞儞僷僋幙偑崌惉偝傟傞丅 恄宱儂儖儌儞丒僆僉僔僩僔儞偱愢柧偟傛偆丅
嘇亅乮侾乯僞儞僷僋幙崌惉奐巒暋崌懱偺宍惉丗 傑偢丄mRNA偲儂儖儈儖儊僠僆僯儞丒tRNA偑丄儕儃僜乕儉乮捈宎20僫僲儊乕僞乕埵偺彫偝側棻偱偡乯偺彫僒僽儐僯僢僩乮恾拞忋懁乯偵寢崌偡傞丅摉慠偺帠側偑傜丄儂儖儈儖儊僠僆僯儞丒tRNA偼mRNA偺奐巒僐僪儞偲憡曗寢崌傪偡傞丅傑偨mRNA偵偼丄奐巒僐僪儞乮AUG乯偺俉偐傜侾俁墫婎傎偳忋棳偵摿暿側墫婎攝楍偑偁傝丄偙傟偲彫僒僽儐僯僢僩偑憡屳嶌梡偡傞偙偲偵傛偭偰丄撉傒庢傝榞偺揔惓側埵抲寛傔偑側偝傟傞丅偙偺忬懺丄偡側傢偪彫僒僽儐僯僢僩亅mRNA亅儂儖儈儖儊僠僆僯儞丒tRNA傪彫僒僽儐僯僢僩奐巒暋崌懱乮initiation complex乯偲屇傇丅忋恾偼僞儞僷僋幙崌惉偑偐側傝恑傫偩忬懺傪帵偟偰偄傞偺偱丄斀墳奐巒帪偵尒傜傟傞彫僒僽儐僯僢僩奐巒暋崌懱傪憐憸偡傞偵偼丄mRNA嵔傪僘儖僘儖乕僢偲塃懁傊堷偄偰壓偝偄丅恾拞儂儖儈儖儊僠僆僯儞偺棯崋偼fMet偱偡丅 偮偓偵丄儕儃僜乕儉偺戝僒僽儐僯僢僩乮恾拞壓懁乯偑嬤偯偄偰偒偰丄彫僒僽儐僯僢僩偲懳岦偡傞埵抲偱椉僒僽儐僯僢僩偼崌懱偟丄儂儖儈儖儊僠僆僯儞丒tRNA偲寢崌偡傞丅偙偺忬懺丄偡側傢偪彫僒僽儐僯僢僩亅mRNA亅儂儖儈儖儊僠僆僯儞丒tRNA亅戝僒僽儐僯僢僩傪奐巒暋崌懱偲屇傇丅偙偺寢壥丄儕儃僜乕儉丄mRNA偦偟偰儂儖儈儖儊僠僆僯儞丒tRNA偺俁幰偑僙僢僩偝傟丄偙偺忬懺傪僞儞僷僋幙崌惉奐巒暋崌懱偺宍惉偲屇傃丄僞儞僷僋幙惗崌惉偺弨旛偑惍偭偨偺偱偁傞丅忋恾偼僞儞僷僋幙崌惉偑偐側傝恑傫偩忬懺傪帵偟偰偄傞偺偱丄斀墳奐巒帪偵尒傜傟傞奐巒暋崌懱傪憐憸偡傞偵偼丄mRNA嵔傪僘儖僘儖乕僢偲塃懁傊堷偄偰壓偝偄丅恾拞儂儖儈儖儊僠僆僯儞偺棯崋偼fMet偱偡丅 嘇亅乮俀乯儁僾僠僪嵔偺怢挿乮僞儞僷僋幙偺崌惉乯偲廔巭丗 戝僒僽儐僯僢僩偲彫僒僽儐僯僢僩偑崌懱偟偨儕儃僜乕儉昞柺偵偼丄傾儈僲巁偑寢崌偟偨tRNA暘巕俀屄偑擖傝崬傔傞孍傒偑偁傞丅恾拞嵍懁偑P乮儁僾僠僕儖乯晹埵偱丄塃懁偑A乮傾儈僲傾僔儖乯晹埵偲屇偽傟傞丅mRNA嵔偼丄偙偺晹埵傪俁僰僋儗僆僠僪乮侾偮偺傾儈僲巁偵懳墳偡傞乯偢偮嵍傊恑傒側偑傜丄僞儞僷僋幙偑崌惉偝傟傞丅 恾拞mRNA偺奐巒僐僪儞乮AUG乯偺塃椬偺僐僪儞乮UGU乯偵懳偟偰丄傾儞僠僐僪儞乮ACA乯傪桳偡傞傾儈僲巁乮Cys丄僔僗僥僀儞乯偺偮偄偨Cys丒tRNA偑傗偭偰偒偰A乮傾儈僲傾僔儖乯晹埵偵寢崌偡傞丅偡傞偲丄P晹埵偺儂儖儈儖儊僠僆僯儞丒tRNA乮P晹埵乯偐傜儂儖儈儖儊僠僆僯儞乮fMet乯偑奜傟偰丄A晹埵偺傾儈僲巁乮Cys丄僔僗僥僀儞乯偲儁僾僠僪寢崌偡傞丅偙偺寢壥丄A晹埵偼fMet丒Cys丒tRNA偲側傝丄P晹埵偼fMet偑庢傟偨tRNA偲側傞丅偟偐偟丄偙偺忬懺偼晄埨掕側偺偱丄A晹埵偺fMet丒Cys丒tRNA偼嵍懁偺P晹埵傊堏傞丅摨帪偵mRNA傕俁僰僋儗僆僠僪乮侾偮偺傾儈僲巁偵懳墳偡傞乯嵍傊恑傓丅傑偨丄P晹埵偵嫃偨tRNA偼宯奜傊曻傝弌偝傟傞丅 偝傜偵丄Tyr丒tRNA偑傗偭偰偒偰A晹埵偵寢崌偡傞丅偡傞偲嵞傃丄摨偠僾儘僙僗偑孞傝曉偝傟偰丄P晹埵偺fMet丒Cys丒tRNA偐傜fMet丒Cys偑奜傟偰丄A晹埵偺Tyr偲儁僾僠僪寢崌偡傞丅偙偺寢壥丄A晹埵偼fMet丒Cys丒Tyr丒tRNA偲側傝丄P晹埵偼fMet丒Cys偑庢傟偨tRNA偲側傞丅偟偐偟丄偙偺忬懺偼晄埨掕側偺偱丄A晹埵偺fMet丒Cys丒Tyr丒tRNA偼嵍懁偺P晹埵傊堏傞丅摨帪偵mRNA傕俁僰僋儗僆僠僪乮侾偮偺傾儈僲巁偵懳墳偡傞乯嵍傊恑傓丅傑偨丄P晹埵偵嫃偨tRNA偼宯奜傊曻傝弌偝傟傞丅偙偺傛偆偵偟偰丄儁僾僠僪嵔偼師乆偲怢傃偰偄偔丅mRNA偺廔巭僐僪儞偵払偡傞偲儁僾僠僪嵔偺怢挿偼巭傑傞丅偙偺帪揰偱丄儁僾僠僪嵔偲tRNA偺寢崌偼愗傝棧偝傟丄儁僾僠僪嵔偼宯奜傊弌偰峴偔丅 僞儞僷僋幙崌惉斀墳偺廔巭偵偼丄mRNA拞偺廔巭僐僪儞乮UAA丄UAG丄UGA乯偺懚嵼偼柍榑偺偙偲丄廔寢場巕乮release factor,RF乯偲偄偆僞儞僷僋幙偑廳梫側栶栚傪壥偨偟偰偄傞丅 埲忋丄mRNA偺俁墫婎偢偮偺墫婎攝楍乮僐僪儞偲屇傇乯偵懳墳偡傞傾儈僲巁偑丄師乆偲楢寢偟偰僞儞僷僋幙乮傾儈僲巁偑偮側偑偭偨崅暘巕暔幙乯偑嶌傜傟傞夁掱傪東栿偲屇傫偱偄傞丅 僐僪儞偺庬椶偼丄墫婎偺庬椶偑係乮A丄U丄G丄C乯側偺偱丄係亊係亊係亖俇係偲側傞丅偙偺偆偪俇侾庬椶偑傾儈僲巁偺庬椶傪寛傔傞偺偵巊傢傟丄俁庬椶偼廔巭傪堄枴偡傞丅僞儞僷僋幙拞偺傾儈僲巁偺庬椶偼俀侽側偺偱丄堚揱埫崋偺僐乕僪昞偵傕尒傜傟傞傛偆偵丄堦偮偺傾儈僲巁偵懳偟偰暋悢偺僐僪儞偑懳墳偟偰偄傞丅 僸僩偺嵶朎偱偼丄悢枩庬椶偺僞儞僷僋幙偑崌惉偝傟傑偡偑丄偦偺峔惉惉暘偱偁傞傾儈僲巁偺庬椶偼嬐偐俀侽庬偱偡丅幹懌偱偡偑丄傾儈僲巁偑偨偔偝傫偮側偑偭偨崅暘巕傪僞儞僷僋幙偲屇傫偱偄傑偡丅 恄宱儂儖儌儞丒僆僉僔僩僔儞丗 俋屄偺傾儈僲巁偑偮側偑偭偰偄傞丄擼壓悅懱偵偁傞恄宱儂儖儌儞偱丄巕媨暯妸嬝偺廂弅嶌梡丄曣擕偺曻弌嶌梡側偳偵娭學偡傞丅 傾儈僲巁攝楍丗 Cys-Tyr-Ile-Gln-Asn-Cys-Pro-Leu-Gly 僐僪儞懳斾昞丗
堚揱埫崋偺僐乕僪昞乮mRNA偺僐僪儞偵懳偡傞傾儈僲巁乯丗
俀侽庬偺傾儈僲巁偲棯崋 僌儕僔儞乮Gly乯丄傾儔僯儞乮Ala乯丄僶儕儞乮Val乯丄儘僀僔儞乮Leu乯丄僀僜儘僀僔儞乮lle乯丄僾儘儕儞乮Pro乯丄僙儕儞乮Ser乯丄僗儗僆僯儞乮Thr乯丄僔僗僥僀儞乮Cys乯丄儊僠僆僯儞乮Met乯丄傾僗僷儔僊儞巁乮Asp乯丄傾僗僷儔僊儞乮Asn乯丄僌儖僞儈儞巁乮Glu乯丄僌儖僞儈儞乮Gln乯丄儕僕儞乮Lys乯丄傾儖僊僯儞乮Arg乯丄僸僗僥傿僨傿儞乮His乯丄僼僃僯儖傾儔僯儞乮Phe乯丄僠儘僔儞乮Tyr乯丄偦偟偰僩儕僾僩僼傽儞乮Trp乯丅 tRNA tRNA暘巕偼丄俈俁偐傜俋俁屄偺僰僋儗僆僠僪乮墫婎偲儕儃乕僗偲儕儞巁偐傜側傞RNA偺婎杮峔惉扨埵乯偱峔惉偝傟傞侾杮嵔乮暘巕検偼俀枩俆侽侽侽偐傜俁枩乯偱丄L帤宆偺棫懱峔憿傪偟偰偄傞丅tRNA暘巕偼侾杮嵔偱偁傞偑丄墫婎偑偍屳偄偵悈慺寢崌傪偟偰偄傞偨傔偵撈摿偺棫懱峔憿傪偲傞丅僀儊乕僕偲偟偰偼丄L帤偺俀偮偺愭抂偺偆偪丄堦曽偵偼摿掕偺傾儈僲巁乮侾庬椶乯偑寢崌偡傞晹埵偑桳傝丄懠曽偵偼mRNA偺摿掕偺墫婎攝楍乮傾儈僲巁偺庬椶傪寛傔傞墫婎俁屄偺慻傒崌傢偣丗僐僪儞偲偄偆乯偲憡曗寢崌傪偡傞墫婎攝楍乮傾儞僠僐僪儞乯偑偁傞丅傑偨侾偮偺tRNA暘巕偼丄侾庬椶偺傾儈僲巁偦偟偰mRNA暘巕偺侾庬椶偺僐僪儞偵偟偐懳墳偱偒側偄偺偱丄tRNA暘巕偺庬椶偼丄傾儈僲巁偺庬椶乮俀侽庬乯偩偗偁傞丅偟偐偟幚嵺偼傕偭偲懡偄偑丅 mRNA DNA偺堚揱忣曬偼墫婎攝楍偱偁傞偑丄僞儞僷僋幙偑崌惉偝傟傞夁掱偱偼丄偄偭偨傫RNA偺墫婎攝楍偵抲偒姺偊傜傟傞丅偙偺RNA傪儊僢僙儞僕儍乕RNA乮mRNA乯偲屇傇丅乷惓妋偵尵偆偲偪傚偭偲堘偆丅DNA偺墫婎攝楍偺揮幨偵傛偭偰嶌傜傟偨mRNA偼丄mRNA偺慜嬱懱偱偁傝丄僀儞僩儘儞乮傾儈僲巁攝楍偵偼娭學柍偄乯偲僄僉僜儞乮戝帠側晹暘乯偲偄偆晹暘偑偮偄偰偄傞丅偙偺晄梫側僀儞僩儘儞偑嶍彍乮僗僾儔僀僔儞僌偲偄偆乯偝傟偰mRNA偲側傞丅乸 mRNA偺墫婎攝楍偼DNA偺俀杮偺嵔偺偆偪堦曽偺墫婎攝楍偲摨偠偱偁傞偑丄T偑U乮僂儔僔儖乯偵曄傢偭偰偄傞丅 儕儃僜乕儉 嵶嬠埲奜丄偡側傢偪恀妀惗暔偺嵶朎偼丄妀枌偵曪傑傟偨妀偲嵶朎幙偑摿挜揑偱偁傞丅嵶朎幙偵偼丄儕儃僜乕儉丄儈僩僐儞僪儕傾丄僑儖僕懱丄庬乆側彫朎懱乮慹柺彫朎懱丄妸柺彫朎懱側偳乯丄塼朎丄嵶朎幙婎幙側偳偑偁傞丅 儕儃僜乕儉偼丄慹柺彫朎懱偺枌柺偵寢崌偟偰偄傞棻巕忬偺RNA偲僞儞僷僋幙偺暋崌懱乮斾棪偼侾丗侾乯偱偁傝丄戝僒僽儐僯僢僩偲彫僒僽儐僯僢僩偱峔惉偝傟傞偑丄僞儞僷僋幙崌惉帪偵偼戝彫偺僒僽儐僯僢僩偵暘棧偡傞丅傑偨僞儞僷僋幙崌惉偼儕儃僜乕儉偺昞柺偱峴傢傟丄侾屄偺儕儃僜乕儉偼侾搙偵侾杮偺億儕儁僾僠僪嵔乮挿偄偺傪僞儞僷僋幙偲偄偆乯傪偮偔傞丅忦審偵傛偭偰偼丄侾侽昩娫偱係侽侽屄偺傾儈僲巁暘巕傪娷傓億儕儁僾僠僪嵔偑崌惉偝傟傞丅 DNA 嵶朎偺捈宎偼10儈僋儘儞掱搙偱偁傝丄偦偺拞怱晹偵偼嵶朎妀偑偁傞丅嵶朎妀偺庡惉暘偼丄巁惈傪帵偡暔幙丄妀巁偱偁傝丄僨僆僉僔儕儃妀巁乮DNA乯偲屇偽傟偰偄傞嵶挿偄暘巕偱丄俀杮偺懳偲側偭偰偄傞丅 俀杮嵔DNA偺棫懱峔憿偼丄1953擭3寧丄悽奅揑偵桳柤側壢妛嶨帍乽Nature乿偵敪昞偝傟傑偟偨丅儚僩僜儞偲僋儕僢僋偺丄俀杮偺嵔偑傛偠傟偁偭偨俀廳傜偣傫峔憿儌僨儖偑偦傟偱偁傞丅 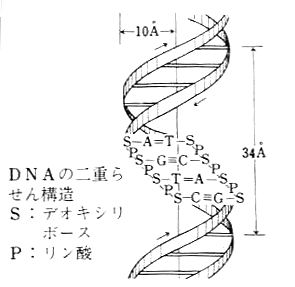 僀儊乕僕揑偵偼丄俀杮偺DNA暘巕偱嶌傜傟偨丄傜偣傫奒抜乮嵍恾偼棟壢擭昞傛傝敳悎乯偱偁傞丅懄偪丄傝傫巁乮P乯偲摐乮僨僆僉僔儕儃乕僗丄S乯偑岎屳偵壔妛寢崌偟偰丄挿偄嵔乮奒抜偺庤悹偵憡摉偡傞乯傪宍惉偡傞丅嵔拞偺奺摐乮僨僆僉僔儕儃乕僗乯偺摿掕偺埵抲偵偼丄侾暘巕偺墫婎暘巕乮A,G,T,C偺係庬椶偁傝乯偑壔妛寢崌傪偟偰偄傞丅堦曽偺嵔偺墫婎暘巕偼丄懠曽偺嵔偺墫婎暘巕偲悈慺寢崌乮壔妛寢崌傛傝寢崌椡偑庛偄偺偑戝愗側億僀儞僩乯傪偟偰墫婎懳乮奒抜偺僗僥僢僾偵憡摉偡傞乯傪偮偔傝丄嵍塃偺嵔偑棧傟側偄傛偆偵堷偒晅偗偰偄傞丅偙偙偱丄悈慺寢崌傪宍惉偡傞墫婎懳偺慻傒崌傢偣偵偼丄乽摿掕偺憡庤偵偺傒寢崌偟偰摿掕偺墫婎懳乮俀庬椶偟偐偁傝傑偣傫乯傪偮偔傞乿偲偄偆婯懃乮憡曗寢崌惈乯偑偁傞丅懄偪丄A乮傾僨僯儞乯偼T乮僠儈儞乯偲丄偦偟偰C乮僔僩僔儞乯偼G乮僌傾僯儞乯偲偺傒墫婎懳傪偮傞丅傑偨丄墫婎懳偼暯柺峔憿傪偟偰偄傞丅 僀儊乕僕揑偵偼丄俀杮偺DNA暘巕偱嶌傜傟偨丄傜偣傫奒抜乮嵍恾偼棟壢擭昞傛傝敳悎乯偱偁傞丅懄偪丄傝傫巁乮P乯偲摐乮僨僆僉僔儕儃乕僗丄S乯偑岎屳偵壔妛寢崌偟偰丄挿偄嵔乮奒抜偺庤悹偵憡摉偡傞乯傪宍惉偡傞丅嵔拞偺奺摐乮僨僆僉僔儕儃乕僗乯偺摿掕偺埵抲偵偼丄侾暘巕偺墫婎暘巕乮A,G,T,C偺係庬椶偁傝乯偑壔妛寢崌傪偟偰偄傞丅堦曽偺嵔偺墫婎暘巕偼丄懠曽偺嵔偺墫婎暘巕偲悈慺寢崌乮壔妛寢崌傛傝寢崌椡偑庛偄偺偑戝愗側億僀儞僩乯傪偟偰墫婎懳乮奒抜偺僗僥僢僾偵憡摉偡傞乯傪偮偔傝丄嵍塃偺嵔偑棧傟側偄傛偆偵堷偒晅偗偰偄傞丅偙偙偱丄悈慺寢崌傪宍惉偡傞墫婎懳偺慻傒崌傢偣偵偼丄乽摿掕偺憡庤偵偺傒寢崌偟偰摿掕偺墫婎懳乮俀庬椶偟偐偁傝傑偣傫乯傪偮偔傞乿偲偄偆婯懃乮憡曗寢崌惈乯偑偁傞丅懄偪丄A乮傾僨僯儞乯偼T乮僠儈儞乯偲丄偦偟偰C乮僔僩僔儞乯偼G乮僌傾僯儞乯偲偺傒墫婎懳傪偮傞丅傑偨丄墫婎懳偼暯柺峔憿傪偟偰偄傞丅偙偺憡曗寢崌惈偺偍偐偘偱丄俀杮偺DNA嵔偺偆偪丄堦曽偺墫婎攝楍偑寛傑傟偽丄懠曽偺墫婎攝楍偑寛傑傞偙偲偲側傞丅懄偪丄DNA偺暋惢帪偵偼丄擇廳傜偣傫偑傎偳偗丄奺乆偺嵔偵懳偟偰憡曗揑側嵔偑崌惉偝傟傞偺偱丄傑偭偨偔摨偠擇廳傜偣傫偑俀慻偱偒傞偙偲偲側傞丅 傑偨丄壔妛寢崌傛傝庛偄悈慺寢崌偺偍堿偱丄揮幨僾儘僙僗偺偲偙傠偱弎傋偨傛偆偵丄RNA億儕儊儔乕僛偵傛偭偰俀杮嵔DNA偑侾杮嵔DNA偲側傞丅 |