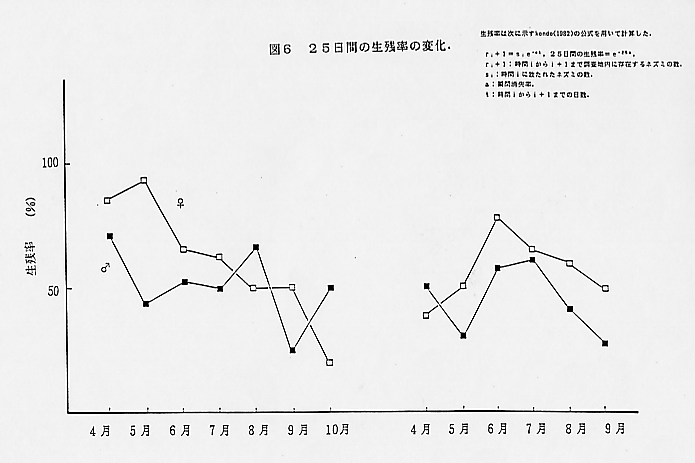
2.生残率の変化
図6は図5での捕獲個体数を用いて、雄と雌の生残率の変化について示している。調査期間の間の日数が異なっているため、各月の生残率はKondo(1982)の公式を用いて計算した。ここでの生残率は25日間の生残率を示す。ここでは成体の生残率についてのみ示してある。雄の生残率の変化についてみてみると、1985年の4月の生残率が高く、その後5月になると生残率が低下する。このレベルは7月まで続き、8月に上昇し、その後9月に(交尾期)急激に低下し、10月に再び高くなる。また生残率の大きな低下は、その前に多くの移入がある時期に続いて起こる。1986年も5月に低下が見られ、その後6月と7月に生残率が上昇する。6月に多くの移入があるが、この時期生残率が上昇している。これは1985年とは違ったパターンである。1985年には8月に生残率の上昇が見られるが、1986年の8月には低下が見られ、9月に生残率は最低になる。1985年にくらべて、1986年では夏の間の生残率が高くなっている。
雌の生残率は雄とは違う変化を示す。1985年は春に生残率が高く、その後段々と低下し、10月(出産・授乳期)に大きく低下する。夏場の生残率はあまり大きな変化は見られない。1986年は4月の生残率が悪く、その後6月まで上昇する。1985年では低下は6月から見られるが、1986年では7月からで、これは6月に多くの新個体が捕獲された割には移出個体が少なかったためである。1985年同様、1986年の夏場も生残率にあまり大きな変化はないが、1985年に比べ、1986年の夏場は雄と同様に高いレベルを示す。また両年とも雄に比べ雌の方が生残率が高く、これは前記の個体数変化のところで見た雄の移入・移出の多いこと、雌で移入・移出があまり多くないことを示す。